
The video may be found at low and high resolution at StudioDaily. If it's at all possible, scroll down a bit and view the high resolution link.
Regardless of the actually story, and there is a story, this is an animation of what's going on inside each of your trillions of cells every moment of every day. You are alive because of this. Glenn and I couldn't help but watch this over and over again this morning. We recognize receptor proteins, microtubules being assembled and disassembled, circular RNAs being propelled from nuclear pores, and translation and insertion into the golgi. There's dramatic exocytosis, peripheral proteins floating on a phospholipid membrane sea and integral proteins hanging from the cell wall above, a lymphocyte squeezing through a capillary wall, and my hands down favorite, the heroic kinesin trundelling along a microtube hauling a massive organelle. And much much more. I haven't had to copy and paste this description from anyone; I recognize everything I'm seeing (although it took awhile, like a good murder mystery). And it's not particularly my expertise, just a bit of familiarity with the things going on. It is that the animation has been so well rendered, although not without a degree of artistic license that I fully applaud.
A lot of people loved the video but have no idea what's going on. You don't have to, to enjoy it, because it's lovely artistry, but I pieced together the story, I think. Wanna know more?
This appears to be the story of the activation of a helper T-cell, although to what extent, I'm still not sure. A teensy bit of background: you have macrophages, a type of white blood cell, that scour your body searching for foreign invaders. When it finds them, it ingests and breaks them down. It then presents the foreign proteins on its surface. Helper T-cells come along and sample it, and when the right one finds it, as in this video, that helper T-cell undergoes changes that allow it to activate and stimulate toward massive reproduction not just any, but precisely the right cytotoxic T-cells and B-cells that produce antibodies, the minute favored among millions. The fight against infection is on.
This video appears to address only the earliest stages - the activation of the helper. (It may be even a smaller story - the story of how a lymphocyte decides to leave the bloodstream and squeeze through the capillary cells.)
I've made screenshots, in order, and have described them below. Click on the thumbnails for a larger view opening into a new page.
 | The video opens with red blood cells streaking past us through an arteriole. The crawling white spheres are lymphocytes, white blood cells. Each can "smell" only one interesting thing. One is about to find his odor, and everything in him will scream "STOP". |
 | Our hero the lymphocyte (bottom) has attached to proteins being present from another cell (above) This could be the Major Histocompatibility Complex, a sort of hand-shaking device by which lymphocytes know not to interact with foreign or inappropriate cells. Or maybe it's just Cell Adhesion Molecules. |
 | A wonderful depiction of membrane proteins floating on lipid rafts on the outside surface of the cell, a phospholipid sea. The membrane proteins are wildly varied, and carry out all sorts of interesting functions. Right now they are translating the word "STOP!", and passing the information on into the cell itself. |
| One of these functions is transduction. A stimulated membrane protein is nestling into another one. This will begin a cascade of events that must take us into the cell's interior. | |
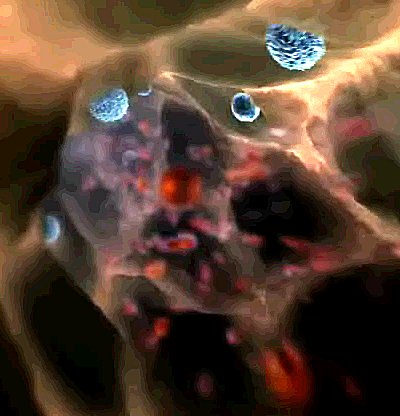
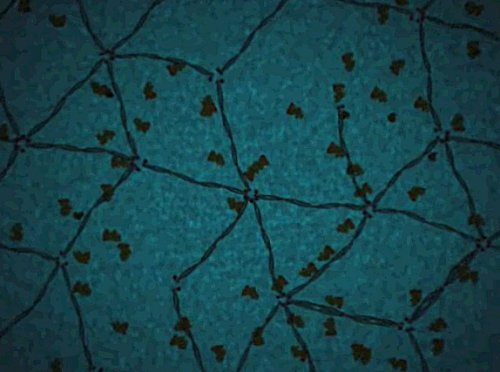 | Neither Glenn nor I can figure out the significance of the pentagonal pattern of reticulation on the cell surface, just before we enter. |
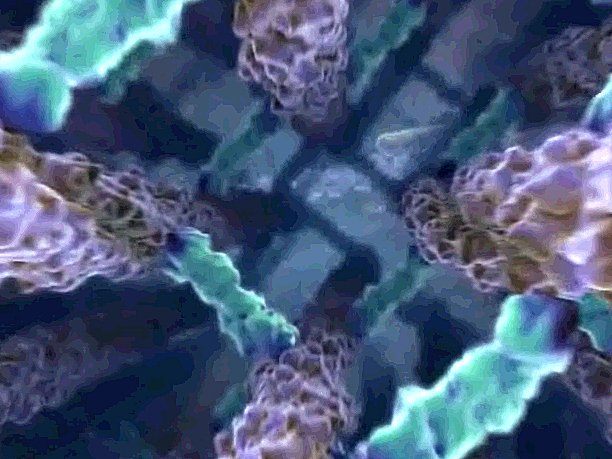 | The descent through the outer architecture into the cell's interior. The next view opens up into an assembling array of microfilaments, part of the cytoskeleton. |
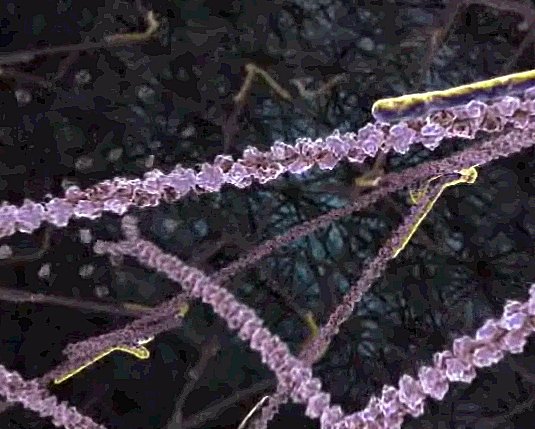 | Microfilaments are one of the three types of cytoskeleton, assembled from smaller building blocks. They are responsible for cell movement and cell shape, as well as simply holding the cell together. These microfilaments are being rapidly constructed in response to the signal received a short time ago. They will brake the cell, and then they will force the cell into a new pattern of movement. |
 | Flurries of new activity. An enzyme has latched onto an actin filament and has snipped it into two pieces. There has to be a reason for this. |
 | A second portion of the cytoskeleton, the microtubules, is being assembled. These are the highways along which the organelles, those tiny parts of a cell, travel and are conveyed. |
 | Microtubules are not only assembled at one end, they are disassembled at the other, giving the impression of a forward direction of movement when in fact there is none. |
 | The motor protein, kinesin. Surely the star of the show, and everyone thinks it's a concoction. "Is this real??" they ask. Yes it's real. It latches onto an organelle with its "arms" and then literally steps along the microtubule path, always placing its "feet" into the groove, one step at a time. This is how the cell moves its parts around. |
 | The enormous organelle the stalwart kinesin guy is carrying is a vesicle filled with proteins destined for the outside, perhaps in response to the signal that the cell had received a while back. The vesicle itself has membrane proteins studding its surface. If you look closely in this sequence, you see hundreds of fellow kinesin proteins hauling their own burdens along the microtubule highways. |
 | Now we get to see the artists' concept of the vast interior of the cell, and in the fuzzy distance, the microtubules radiate outward from the centrosome - Microtubule Organizing Center, the MTOC, dominating the proceedings. |
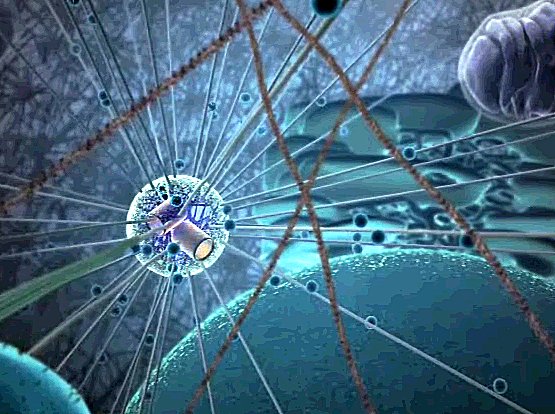 | The focus shifts toward the nucleus in the foreground, and the MTOC with its star pattern of microtube highways radiating from the cylindrical centrioles to all parts of the cell. These are controlling the events of the moment. Now we're going to see what the nucleus is doing. |
| Strings of messenger RNA are shooting up out of the nucleus through the nuclear pores. These include the new programming instructions that will produce the proteins that will cause this cell to change its behavior. | |
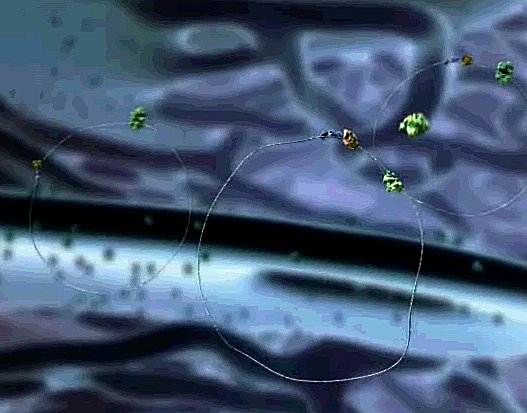 | The RNAs drift above the nucleus and circularize, touching G-caps to polyA tails. The green ribosomes approach, and latch on. They slide around the glistening circle of mRNA, quickly extruding the protein, and then everyone falls off the messenger strand. |
| This ribosome is busily spinning a protein, moving quickly along the messenger RNA strand, translating it, and adding each new amino acid one by one. The protein folds into its proper shape as you watch. | |
 | Two newly synthesized proteins latch onto one another. A fit! They drift across that manatee-like body, the mitochondrion, from which all energy for all this activity comes. |
| Translation is occuring at the surface of the Endoplasmic Reticulum, too. Ribosomes threaded with mRNA sit down atop an ER pore, and spin their protein right down into the lumen. This protein will whisked off to the golgi apparatus, to be sorted and sent on to its ultimate destination, probably ending up in one of the vesicles being hauled by our kinesin motor protein friends. | |
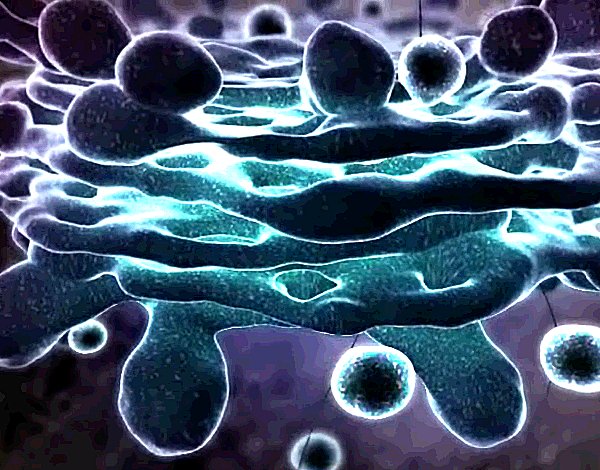 | Speak of the devil! The great, slowly rotating, pulsating galaxy is the golgi apparatus that so bored you in school. This is where the above proteins are headed, to be processed, modified, marked and put into their appropriate vesicles. Delivery to any one of a multitude of destinations is by kinesin express. |
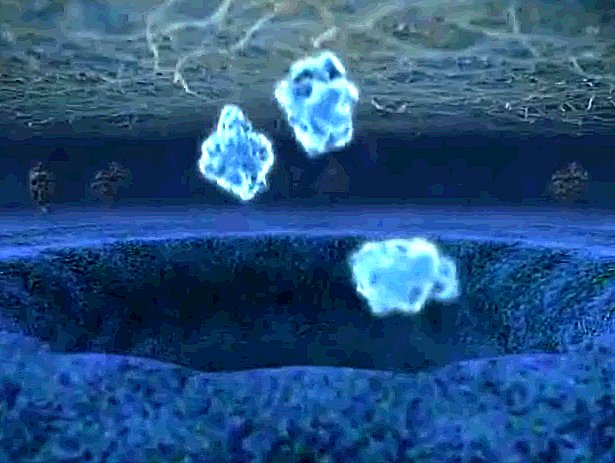 | Now we see the fate of the proteins we've seen being made. A huge bubble emerges from the interior of the cell, and bursts, scattering some proteins out into the void. Perhaps these are interferon, or cytokines, and they themselves are messages to other cells that will start a cascade of changes within them. This is exocytosis. |
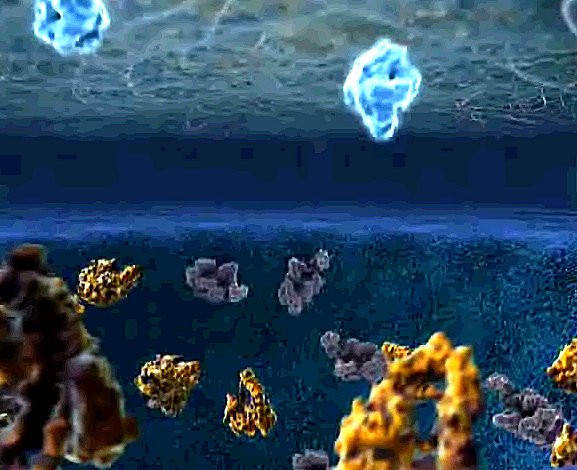 | Not all proteins are released into the void. Some stick to the outer membrane surface. These are receptor proteins, transducer proteins, channel proteins, and much much more. |
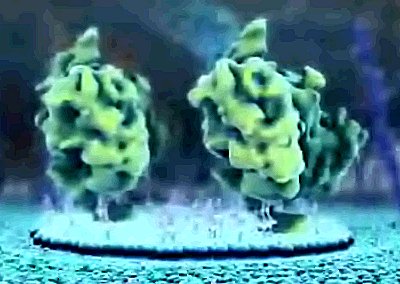
 | A lipid raft gathers under a group of the newly arrived proteins. |
 | They rise up, and begin to unfold, stretching above the scaffolding hovering above the cell's surface. |
 | Their binding sites are now above the scaffolding and they are prepared to latch onto anything their sticky hands can grasp. Which is to say, only one thing, that which they were made for. |
 | And what do they grasp? Apparently just the surface of the arteriole. The lymphocyte rolls to a halt. (And this is why I think maybe this has to do more with cell adhesion than T-cell receptors, but it really doesn't matter too much.) |
 | Having stopped its mindless forward motion, the lymphocyte now squeezes out of the bloodstream and into the cellular interstitial spaces, looking for trouble. |